


The essential difference here is that the 'fruity' portion of the juniper berry developed from the fusion of the scales surrounding the bare seed; that of the others, from a flower's ovary. But the seeds of all three contain one or more cotyledons.
Below I list traits associated with monocots, but each of these all must be qualified by "in general." They may not necessarily hold for any individual family or genus.

Four dicotyledons shown are Parkinsonia aculeata (retama), Sophora secundiflora (Texas mountain laurel), Chilopsis linearis (desert willow) and Arbutus xalapensis (Texas madrone). The cotyledons of S. secundiflora remain below the surface (are 'hypogeous'). Those of C. linearis are deeply cleft, almost appearing as four. Note that in all of these images the true vegetative leaves are above the cotyledons and have a different shape or texture. But if the cotyledons rise above the surface, chlorophyll is produced, with photosynthesis much as with true leaves.




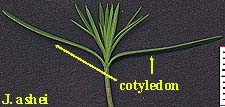
Although nearly everyone who has raised plants from seed is familiar with the pairs of sprout leaves that accompany the germination of dicots. The cotyledon of a monocot is not so easily observed, and not just because it remains below the surface (as with dicot Texas mountain laurel). The literature on monocot cotyledons is generally uninformative if not misleading. The University of Kentucky's College of Agriculture site Plant Physiology I, Lecture 8: Seed and fruit development provides an explanation that seems to account for the difficulties I have encountered searching for the cotyledons of monocots:
The single cotyledon of the monocots (the scutellum Fig. 1, 3A) is usually of a secretory and absorptive nature, never exiting the seed proper, even after germination is complete. It abscises from the seedling and is shed along with the exhausted endosperm and testa upon the completion of seedling establishment. It rarely contains substantial amounts of stored reserves being associated with endospermic seeds (seeds in which the major storage organ is an endosperm).In conjunction with grasses, we frequently encounter statements such as that in F. G. Gould, Grass Systematics (1968; p. 362):
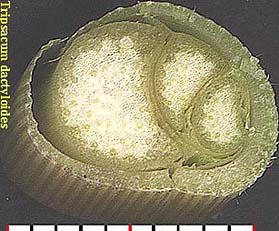
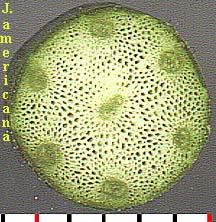
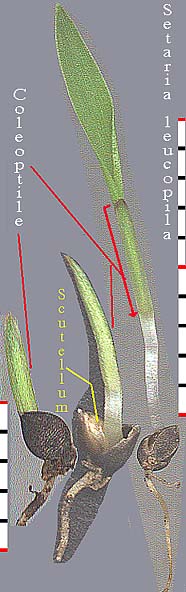
Scutellum. Haustorial tissue of the embryo located between the endosperm and the plumule-radicle axis of the embryo; considered by some to be the grass cotyledon. [emphasis by Bob Harms]Two series of grass germination images, for Sporobolus clandestinus (rough dropseed) and Setaria leucopila (yellow foxtail), are given below. The scutellum, visible with the second set, has been labeled. Shown in both is the 'rapid elongation of the embryonic shoot and emergence of the coleoptile' (Gould, p. 29), a sheath from which the true leaf can be seen emerging.

Although no monocot has two cotyledons, in the rudimentary embryo of orchids, consisting of only a few cells, the process of simplification seems to have reached a stage of no cotyledon at all.
In sum, the monocot cotyledon appears to be a phylogenetic issue likely to elude the amateur botanist hoping to observe it.

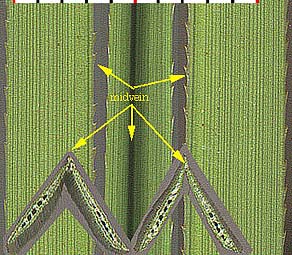

Some monocots are said to have netted veins, such as Smilax (Correll and Johnston 1970, p. 409), but in both Smilax tamnoides (bristly greenbrier) and Potamogeton illinoensis (Illinois pondweed), shown below, one senses that the reticulation is superimposed on a primary parallel vein pattern.







The lily
Schoenocaulon texanum (green lily) has 2 whorls of 3 stamens (left), 3 styles (right), and 2 whorls of essentially identical sepals and petals.


Iris hexagona (Dixie iris) reveals its flower parts to be in multiple whorls of 3.

The monocot Potamogeton illinoensis (Illinois pondweed) deviates from the expected pattern with 4 sepal-like segments, 4 stamens.

Typical among the dicots are Rorippa nasturtium-aquaticum (watercress) with 4s, and Gilia rigidula (bluebowls), here dissected, with 5s.


To illustrate an exceptional dicot, a tripartite floral structure is the rule with Aristolochia serpentaria (Virginia dutchmanspipe), dissected to expose its 6 stamens and 3-lobed style