To Berberis Home
A Short History of the Distinctive Role of Stamen Filaments in Berberis
by Bob Harms
With Focus on Selected North American 'Mahonias'
0. Early botanical sources.
Early botanical sources cited by Linné and later authors make no mention of stamens.
Caspar Bauhin 1623, in his Pinax Theatri Botanici (p. 454) lists three Berberis species and descriptions from Theoprastes and other ancient herbals. Of these only two are recognized by Linné 1753, B. vulgaris and B. cretica (pp. 330-331).

Josephi Pitton Tournefort 1700 (p. 614; Tab. 385) makes
no mention of stamens in his short description of Berberis.
1. Carl Linné — Carl Linnaeus
Linnaeus 1737, p. 94, ¶267:
STAM. Filamenta sex, erecta, compressa. Antherae bifidae, filamentis adnatae.
STAMENS: Filaments six, erect, flattened. Anthers bifid, adnate to the filaments.
Linnaeus (ed. 6) 1764, p. 175, ¶442:
STAM. Filamenta sex, erecta, compressa, obtusa. Antherae duae, filamentoruum apici utrinque adnatae.
STAMENS: Filaments six, erect, flattened, obtuse. Anthers two, adnate on each side to the apex of the filaments.
2. Discovery of new American (and Asian) species in the early 19th century.
Exploration of the American West, especially the Lewis and Clark Expedition (1804-1806) led to discovery of new forms of Berberis with odd-pinnate leaves, thornless stems, and bidentate filaments (cf. the Pursh drawings below).
Mariano Lagasca y Segura 1803 listed the first such new species, B. pinnata, from plants grown in Madrid from seed collected on the American West Coast. The author reference "Lag., Elench. hort. Madr. (1803) 6" leads only to page with two Berberis species listed and no other description - beyond the fact that the name itself means 'Berberis with pinnate leaves.'

Frederick Traugot Pursh 1814 (pp. 219-220, Tab. 4, 5) added two new species from the Lewis and Clark Expedition - B. Aquifolium and B. nervosa
(plus the Berberis proper B. canadensis from the southeastern states) - the first of these described as having:
Filamenta 6. corollae breviora, crassa, medio bidentata: dentibus oppositis.
Filaments 6. Shorter than the corolla, thick, bidentate in the middle: with opposite teeth.

Pursh Tab. 4: B. aquifolium stamen
Pursh's Tab. 5, labeled "Berberis nervosa," has a very small drawing of a petal with stamen which appears to be toothed (enlarged image on the right has been edited).

Pursh Tab. 5: petal with stamen
But as has been noted by several authors (e.g., Hooker 1829; C.G. and J. U. Lloyd in their pamphlet
"Berberidaceae," 1878), although the leaves are from B. nervosa - with unappendaged filaments, the flowers of Tab. 5 belong to B. aquifolium. This may well have been the source of the feature 'bidentate teeth' as a genus character of Mahonia, persisting until the end of the 19th century (cf. Gray 1849 and 1895, Howell 1903) — even by Hooker (cf. below), who noted the mixup in the Pursh drawing.
Thomas Nuttall 1818 introduced a new genus, Mahonia, for the new forms of Berberis. His description of the genus is given as (pp. 211-212, ¶ 307):
MAHONIA.
Calix 6-leaved, unequal. Petals 6. Nectariferous glands none. Filaments irritable, each filiformly bidentate; anthers (as in Berberis) growing to the filaments; cells opening by so many vertical elastic valves. Berry many-seeded.
Suffruticose plants with pinnated leaves and terminal racemes; berries purple.
SPECIES. 1. M. Aquifolium. ... Stamina, filaments linear, flat, about the length of the anthers, with a short filiform process on either side of each, arising from the base of the anther; valves of the anthers erect, conspicuous; cells margined on one side....
2. nervosa. ... [no mention of stamens/filaments] ... - Berberis nervosa. Pursh, 1. p. 219. t. 5.
A third species of this genus is indicated by Mr Pursh as indigenous to ... Napaul in India ....
Nuttall's mention of 'irritable filaments' - i.e., filaments responding to touch by abruptly moving toward the pistil - would seem to indicate an attempt to put Mahonia on a par with Berberis, as this feature was already known for Berberis proper. The absence of nectariferous glands is actually not typical of the Mahonia species, but the description - as with many other erroneous Berberis descriptions - could have stemmed from working with dried specimens, in which case the glands would have been inconspicuous.
De Candolle 1818 (pp. 4-22) recognized Mahonia alongside Berberis and made the difference in filament structure explicit (p. 19):
Genus Berberidi proximum, satis ut videtur distinctum filamentis dentatis, petalis eglandulosis et habitu, omninò foliorum structura confirmans doctrinam de Berberidejam suprà expositam, et inter Berberidem et Nandinam ferè medium.
The closest genus to Berberis, sufficiently distinct when seen with dentate filaments, petals without glands and manner of growth, with the structure of the leaves altogether confirming the features of Berberis set forth above; and nearly midway between Berberis and Nandina.
thus (p. 4):
I. Berberis.
Petala 6, ungue intùs biglanduloso. Stam. 6, filamentis edentulis, antherarum loculis discretis. ...
Petals 6, with two glands on the inside. Stamens 6, filaments without teeth, with separate anther cells.
contrasted with (p. 18):
II. Mahonia.
Berberidis sp. LAG. PURSH. — Mahonia NUTT. gen am. t. n. 307. — Odostemon RAF. amer monthl. mag. 1819. p. 192. [Cf. Odostemon below.]
Petala 6, intùs eglandulosa, sed basi tamen nectarifera (ex Nutt.). Stam. 6, filamentis apice utrinquè 1-dentatis.
Petals 6, without glands on the inside, but nonetheless with a nectariferous base (according to Nutt.). Stamens 6, one toothed on each side of the filament apex.
DeCandolle lists 4 (actually 3) American Mahonia species: M. fasiscularis (= B. pinnata of Lagasca), M. aquifolium, M. nervosa, and M. glumaceae (= M nervosa); plus 2 Asian species M. Napaulensis and with a heading "Species non satis nota" (
Species not sufficiently studied), M. Japonica.
Bonpland, Humboldt, Kunth 1821 (pp. 69-71) apparently still weren't aware of the newly discovered species of Pursh and Nuttall. And although they have Lagasca's B. pinnata, their descriptions make no mention of filament teeth. Their key (p. 69):
1.) Folia simplicia, fasciculata [leaves simple, fasciculate]
[=> 4 species of Berberis proper]
2.) Folia pinnata [leaves pinnately compound; => B. pinnata, no author given]
And their only stamen drawing is of 'toothless' B. conferta.
William Jackson Hooker 1829 (pp. 28-29) describes only North American species and rejects the Berberis—Mahonia split (p. 29):
The genus Mahonia cannot be retained. The teeth of the filaments which were supposed mainly to characterize it, I have shown to exist equally in B. heterophylla ... and the glands are present on the petals....
Even though Hooker calls attention (p. 29) to the mixup of B. nervosa and B. aquifolium in Pursh's Table 5 (cf. Pursh above), and attributes DeCandolle's M. glumacea—M. nervosa confusion to it, he failed to realize that B. nervosa lacks filament teeth. His argument that a Berberis proper species [
heterophylla] also has filament teeth would, of course, have been strengthened by knowledge of a Mahonia-type Berberis that lacked them.
Hooker's key is based on leaf structure alone.
1. BERBERIS. Linn.
... Stam. eduntula (vel dentibus 2 instructa.)
... Stamens without teeth (or fitted with 2 teeth).
* foliis simplicibus. [with simple leaves]
1. B. vulgaris ... Linn. Sp. Pl. p. 472 [identified with Pursh's B. canadensis];
** foliis impari-pinnatis. (MAHONIA. Nutt. DC.) [with odd-pinnately compound leaves]
2. B. pinnata ...
... filamentis bidentatis.[with bidentate filaments]
3. B. Aquifolium ;
... filamentis bidentatis.[with bidentate filaments]
4. B. nervosa ;
... filamentis 2 dentatis. [with 2-toothed filaments]
John Torrey & Asa Gray 1838 (pp. 49-50) do not recognize the genus Mahonia, and although they don't mention filaments in their key for Berberis proper (with two species: B. vulgaris and B. canadensis), filaments are 'somewhat' distinctive in their key for the three 'Mahonias':
§2 ... filaments usually 2-toothed at the summit: Mahonia, Nutt.
3. B. aquifolium ... filaments 2-toothed
4. B. pinnata [nothing about filaments]
5. B. nervosa ... filaments 2-toothed
The key's qualification 'usually' is not explained; and they continue the faulty description of B. nervosa as '2-toothed'; cf. Pursh above; but contrast Gray 1895, p. 70.
3. Odostemon Raf. — a genus founded on dentate stamens.
Odostemon, from Greek οδ
óυσ
tooth +
στημα
stamen, was chosen for the new genus of the Berberis complex by Constantine Samuel Rafinesque in his "Florula missurica" (1817 - which, to my knowledge, was never published), at essentially the same time as Nuttall was publishing his Mahonia genus. "Odostemon Raf." synonyms have subsequently been listed for the two Mahonia species known to Rafinesque, on the basis of his 1818 review of Pursh and his 1819 review of Nuttall (e.g., Gray 1849, p. 79, "ODOSTEMON, Raf. in Amer. Month. Mag. 1819, p. 192"). Apparently the etymology of the name was considered sufficient, for his published reviews provide no discussion or description, just the names:

Total Odostemon/Berberis comment in Rafinesque's 1818 review of Pursh (¶50;).
And in his 1819 review of Nuttall, he adds an attack on the Philadelphia nurseryman Bernard McMahon, for whom Nuttall had named the genus — and who had been asked by Thomas Jefferson to attempt to cultivate the seeds collected by Lewis & Clark in 1807.

Odostemon/Mahonia in Rafinesque's 1919 review of Nuttall (¶56;).
Continued sporadic use of Odostemon by later Botanists as recently as 1910 (Abrams), 1922 (Standley) and 1941 (Tidestrom & Kittell) have resulted in a literature rich with Odostemon synonyms. The florae of these authors present no justification of their selection of this name, no longer distinctive for the genus as a whole, nor do their descriptions provide any information about stamen morphology.
4. Conflicting filament descriptions for three new species of the American Southwest: B. trifoliolata (1841), B. Fremontii & B. haematocarpa (1858, 1898)
a. B. trifoliolata
In 1841
Moise Etienne Moricand published the discovery of a new species, B. trifoliolata, between Laredo and San Antonio (Bejar) (pp. 113-114, tab. 69), with a description of its stamens:
... "Les étamines sont opposées aux pétales, édentées, le filet élargi, l'anthère adnée biloculaire s'ouvrant par une valve blanchâtre qui se roule sur elle-même de la bas au sommet."
The stamens are opposite the petals, without teeth, the filament is enlarged, the anther adnate, bilocular, opening by a whitish valve which rolls up from the base to the summit.

Moricand's stamen drawing scanned from a poor quality
microfiche page (on the left)
with an enhancement
made by connecting dots (right).
This new Berberis with palmately trifoliate leaves and toothless filaments forced a revision of the Berberis complex that should have occurred earlier with B. nervosa.
Gray 1849 (p. 80) both modified the distinctive filament tooth feature of the Mahonia group with the addition of 'salient' and posited a new subgenus for B. trifoliolata:
To the two recognized subgenera [of Berberis] I may here add a third.
§1 BERBERIS proper. — Filaments usually inappendiculate....
§2 TRILICINA. — Filaments inappendiculate. .... (B. trifoliolata, Moric.)
§3 MAHONIA, Nutt. — Filaments appendiculate with two salient teeth at the apex....
Why Gray might have felt obliged to add 'salient' is not evident, since B. nervosa could only be included in his revised Mahonia subgenus on the (erroneous) assumption that its filaments had 'salient teeth.'
Adolf Scheele 1848 (pp. 591-592) quite independently also discovered this new species in the hills at New Braunfels, Texas ("an Felsen bei Neubraunfels"), and assigned it the name B. ilicifolia. But Scheele's plant apparently had appendiculate filaments.
Stamina 6 sepalis opposita, filamenta apice bidentata.
Stamens 6 opposite the petals, filaments with two teeth at the summit.
Since B. ilicifolia - holly-leafed, a rather obvious name for a barberry, already existed among Berberis proper, Scheele immediately changed it in 1849 (p. 154) to B. roemeriana.
With the exception of filament teeth, Scheele's new species proved to be identical to Moricand's B. trifoliolata, and his names were recognized as synonyms by Gray 1895, without discussion of the filament teeth conflict. Fedde 1901 declared the dentate filaments to be a mistake "Die von Scheele l. c. 592 angeführten 'filamenta apice bidentata' habe ich ebenso wie DIPPEL und KÖHNE nicht gesehen. Es muss dies wohl auf einem Irrtum SCHEELE's beruhen" (free translation: Since Dippel, Köhne and I haven't seen the toothless filaments alleged by Scheele, this must be based on an error by Scheele.)
My own recent sampling of B. trifoliolata flowers in Travis and Hays Counties (from 171 plants) shows that filament teeth are uncommon, but do occur in varying degrees. The B. trifoliolata stamens below illustrate the range of filament types (with clearly toothed at the right and the common (c. 60-68% of plants) toothless types at the left). The degree of swelling at the junction of the connective and the filament also varies a great deal. In the image below no two stamens are from the same plant; filament appendages do not vary significantly on individual plants.

B. trifoliolata stamen types
Ahrendt's 1961 monograph on Berberis attempts to clarify the standards for deciding whether a filament is toothed (p.362, fig. 10):
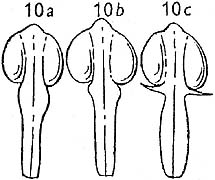
"Fig. 10. Apex of filaments: a, slightly swollen, not dentate...;
b, shortly dentate...; c, conspicuously lanceolate-dentate...."
b. B. Fremontii and B. haematocarpa
Torrey's 1858 (pp. 30-31) survey of the flora along the U.S. – Mexico boundary added a species description for B. Fremontii, said to have been first discovered by Colonel John Charles Fremont in 1844 in southern Utah, noting:
Filaments inappendiculate ... It is nearly allied to B. trifoliolata, but differs in the number of leaflets, longer racemes, and blue berries.
Following Gray's 1849 tripartite subclassification, Torrey assigned it to the (inappendiculate) subgenus Trilicina. Although Gray 1895 recognized Torrey's new species, he rejected the filament description (p. 69):
char. filaments inappendiculate incorrect
and placed it within a group characterized as having "filaments with a pair of divergent or recurved teeth near the apex" (p. 68).
E. O. Wooten 1898 (pp. 304-306) first documented a closely related New Mexico species B. haematocarpa, and resolved the apparent Torrey—Gray conflict (pp. 305-306):
Dr. Torrey had what I take to be this plant [B. haematocarpa] before him on several occasions and confused it with what he finally described as B. Fremontii....B. Fremontii is correctly characterized in ... (Gray 1895, p. 69) ... the filaments are noticeably appendiculate. In B. haematocarpa the flowers are smaller, the appendages to the filaments hardly noticeable....
The combination of inappendiculate filaments with dark blue berries (in contrast with the blood red berries of B. haematocarpa) leaves little doubt that Wooten's interpretation is correct. The closeness of the two species was noted by Fedde 1901 (p. 101):
Die Pflanze [M. haematocarpa] ist ganz nahe verwandt mit M. Fremontii. Man könnte sie vielleicht auch als eine Abart letzterer beschreiben. ... Die Hauptuntershied besteht endlich in der Färbung der Beeren....
The plant is quite closely related to M. Fremontii. One might even perhaps describe it as a variety of the latter. ... The main difference after all is in the color of the berries....
5. A rare Texas endemic, B. swaseyi.
Samuel Botsford Buckley 1870 (pp. 14-15), Texas botanist and geologist, published details of B. swaseyi, discovered in Hays County (Central Texas), on the Pedernales River. [In Young 1873 this is given as "Western Texas on the Perdinales River."] The species was named for H. A. Swasey, editor of Southern Horticulturist, where the description was first published. Buckley did not observe flowers, nor did
E. N. Plank, who reported seeing fruits on a trip to the area in 1893.
John Coulter 1891 presents a short description based on Buckley, with no indication of floral morphology beyond placing it in his key (p. 10) under
"**Filaments without appendages "
along with B. trifoliolata and B. Fremonti [sic.].
Gray 1895, citing only Buckley, Young, Plank and Coulter - none of whom had seen its flowers - places this species in his key under a group with:
"** ...filaments with a pair of divergent or recurved teeth near the apex"
Fedde's 1901 monograph is not so bold as to venture a guess as to the state of B. Swaseyi's filaments, nor does Ahrendt 1961, although he places it in a group specified as '
stamens dentate (where known).'
As with B. trifoliolata above, my survey of some 60 B. swaseyi bushes revealed a wide variety of lengths and forms for the filament appendages, shown in the image below. These range from nearly inappendiculate (on the far right) to broad, long, flexuous appendages on the left. The three in the center, with relatively terete projections represent the norm (67%). As above, no two stamens are from the same bush; stamens were not found to vary significantly on a given bush. And I have not included results from B. trifoliolata X B. swaseyi hybrids — to the extent that these can be recognized independent of stamen morphology. Comparison of the two sets of images reveals an overlapping continuum from inappendiculate to saliently appendiculate. (To view the two sets of images side by side.)

Gray 1895 (pp. 67-70) provided a major revision of Berberis which clearly had great influence on the treatment of the genus in 20th century American florae. His key recognizes two subgenera, Berberis proper and Mahonia, the former with "filaments toothless." The key for Mahonias has three major subgroups based on foliage, bud scales, filaments and fruits. For filaments we find:
* ... filaments toothless [=> B. trifoliolata]
** ... filaments with a pair of divergent or recurved teeth near the apex
[=> seven species known at the time]
*** ... filaments toothless [=> B. nervosa,
which, of course, differs in all other features from B. trifoliolata]
6. Detailed Studies for Berberis & Mahonia
Alfred Usteri 1899 (pp. 77-95) gives a comprehensive classification of the world's Berberis proper (
Euberberis) species. By the end of the century it had become clear that the only feature separating the two sections of Berberis was in their stem (with or without spines representing earlier leaves) and leaf structure (simple or odd-pinnate). Usteri felt that (p. 82):
In der That darf das einzige, konstante Unterscheidungsmerkmal: Die Fiederung der Blätter, nicht zu einer Gattungsbildung Anlass geben.
In point of fact the one constant distinctive feature: compound pinnate leaves, does not allow us to justify the creation of a new genus.
As with earlier Euberberis treatments Usteri recognizes the presence or absence of filament teeth, and uses this as a major distinctive feature in his key, structured as follows (pp. 87-94):
A. Fruit black. ...
I. Filaments always without teeth below the anthers ...
II. Filaments with 2 teeth below the anthers or toothless...
a) Filaments with 2 teeth (in the case of B. buxifolia these may also be absent). Style absent.
b) Filaments without teeth. Style present or absent.
B. Fruit red. ...
[Only one species is marked with filament teeth in this group.]
Friedrich Fedde's 1901 (pp. 30-133) monograph presents a detailed morphological and anatomical analysis of the Mahonia group, and in considering essentially the same data as Usteri (cf. above) he arrives at the opposite conclusion concerning genus status for Mahonia (p. 68):
Das enizig wirklich durchgreifende Merkmal von Mahonia gegenüber Berberis ist also der Besitz von gefiederten Blättern, ein Merkmal, das ich schon ganz für sich allein für geeignet halte, der Gattung Mahonia eine Daseinsberechtigung zu geben.
The only truly decisive feature of Mahonia in contrast with Berberis is thus its pinnate leaf structure, a feature that I consider appropriate quite completely in itself, to entitle the existence of the genus Mahonia.
Although his Latin descriptions use only the term 'tooth' (dens, dent-, edentat-), Fedde's German generally uses 'appendage' (Anhängsel), perhaps a recognition of the inadequacy of the traditional 'tooth' to encompass the wide variety of appendage forms that had been observed.
He presents the first somewhat more detailed sketch of the filament appendages (p. 47)
In den meisten Fällen besitzen die Filamente am oberen Ende dicht unterhalb der Antheren Anhängsel von meist lanzettlich-dreieckiger (M. Aquifolium, repens ...) oder abgerundeter Form (M. Fremontii u. haematocarpa), die bei einzelnen Arten nur als äusserst kleine Vorsprünge bemerkbar sind (M. Fortunei, M. Hartwegi ...). Diese Anhängsel fehlen bei den meisten Longibracteatae, nur bei M. Fortunei sind sie in winzig kleiner Form vorhanden. Ferner fehlen sie noch bei M. trifoliolata und ximapana. Das Vorhandsein oder Fehlen dieser Anhängsel ist für die Unterscheidung der Arten von einem gewissen Wert. Derartige Anhängsel scheinen an den Staubgefässen von Berberis ziemlich selten zu sein.
In most cases the filaments possess appendages of mostly lanceolate-triangular ((M. Aquifolium, repens ...) or rounded (M. Fremontii ssp. haematocarpa) form on the upper end just below the anthers.
With some species these are only noticeable as extremely small projections (M. Fortunei, M. Hartwegi ...).
These appendages are lacking in most of the Longibracteatae, only with M. Fortunei are they present in minutely small form. Moreover they are also absent with M. trifoliolata and ximapana. the presence or absence of this appendage has some value for distinguishing the species. These sorts of appendage seems to be relatively uncommon with the filaments of Berberis.
Diese Anhängsel halte ich für Ligulargebilde, da sie nur aus parenchymatischen Zellen bestehen und keine Spur von Gefässen enthalten. Mit dieser Ansicht im Widerspruch steht die Ansicht CITERNE's ... der diese Zähnchen fur Nebenblattbildungen hält.
I consider these appendages to be a ligular formation, since they consist solely of parenchyma cells and contain no trace of vascular tissue. At variance with this opinion is Citerne's opinion, who considers these little teeth to be stipular formations.
Fedde's Latin teeth description for the Mahonia genus doesn't entirely match his descriptions of individual species ( p. 75):
staminibus 6 duobus cyclis dispositis, antheris muticis valvulis 2 sursum dehiscentibus, duobus dentibus triangularibus infra antheras praeditis rarissime nullis;
with 6 stamens arranged in whorls, with 2 muticous valved anthers opening at the top,
provided with two triangular teeth below the anthers, rarely none
Contrast:
[M. repens, p. 79]
staminibus dentibus duobus triangularibus infra antheras praeditis
stamens provided with two triangular teeth below the anthers
[M. pumila, p. 82]
staminibus dentibus duobus bilobis infra antheras praeditis
stamens provided with two bilobed teeth below the anthers
[M. Fremontii, p. 99]
staminibus dentibus duobus subrotundis parvis infra antheras praeditis
stamens provided with two small nearly rounded teeth below the anthers
[M. haematocarpa, p. 100]
staminibus petala fere aequantibus, infra antheras dentibus duobus rotundatis minimis praeditis;
stamens usually equal the petals, endowed with two minimal rounded teeth below the anthers
[M. nervosa, p. 116]
staminibus edentatis
stamens without teeth
[M. japonica (= M. bealii), p. 118]
staminibus edentatis, filamentis antheras circiter duplo superantibus
stamens without teeth, filaments twice as long as the anthers
It should be clear that even working with dried specimens Fedde must have sensed the descriptive inadequacy of 'toothed' vs. 'toothless.' The image below, contrasts five stamen types, three without teeth and two with; from left to right (1) B. nervosa, (2) B. bealii, (3) B. trifoliolata, (4) B. repens, (5) B. acquifolium. The teeth are not triangular, the toothless filaments and the connectives have quite different forms.
(The left most and right most images are edited from originals by Melinda Denton at the U. of Washington Botany 113 Berberidaceae page. The others are my own.)
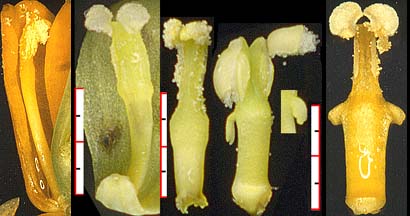
Fedde's Mahonia key (pp. 75-79) utilizes filament structure to distinguish only two species, 3.B.II.a.1: M. paniculata (toothed) and 3.B.II.a.2: M. hartwegii. This, of course, permitted placing of some species, e.g. M. swaseyi, in the absence of filament data without having to guess.
L. W. A. Ahrendt 1961 (pp. 1-410), a relatively recent major monograph on the world's Berberis species, maintains Mahonia as a distinct genus. Although Ahrendt views stamen morphology as significant for diagnostic purposes (pp. 17-18), his treatment is surprisingly simplistic given the published sources from the Western Hemisphere that he accessed, rarely providing detail beyond "dentate" and "edentate." Somewhat indicative of his dated knowledge of American species is his statement (p. 18):
Only three American Mahonias are recorded with edentate stamens, namely M. hartwegii, M. arguta and M. trifoliolata.
which may have been written prior to his description of M. nervosa as "edentate" (p. 329). Worthy of note is the fact that almost 100 years after the discovery of B. swaseyi there was still no botanical description for its flowers/stamens - he bemoans the fact that a 1937 cultivated plant did not flower.
Interestingly, Ahrendt doesn't use the word 'filament' — 'connective' is found. And as a new taxonomic stamen feature he present a typology of stamen apex form, carefully noted for those species he has examined.
In conclusion it must be said that although of no special interest for filament tooth morphology, Ahrendt's monograph contains a wealth of data and classification.
7. 20th Century Floras of the American West
(including The Flora of North America)
For the most part floras of the past century primarily focused on the plants of a region or state, with minimal descriptions beyond features necessary to key out known species.
From a geographical perspective only Pacific Coast floras mentioned filament teeth, and with few exceptions the filament descriptions varied little, if at all, from those given by Torrey and Gray. For example, Torrey & Gray 1838 have:
filaments usually 2-toothed at the summit
compare Howell 1897, 1903
filaments usually 2-toothed at the summit
Again, Gray's 1895 key has
filaments with a pair of divergent or recurved teeth near the apex
which seems to have led to the teeth of virtually all species being described as
recurved.
Compare Jepson 1922, 1925; Abrams 1944; Munz & Keck 1959:
filaments with a pair of recurved teeth near the apex.
and
Whittemore's 1997 FNA paraphrases:
distal end of each anther filament with pair of recurved teeth
anther filaments with(out) distal pair of recurved lateral teeth (repeated for 14 species)
anther filaments distally with pair of recurved teeth
B. haematocarpa (cf. above) is seen with recurved teeth for Munz & Keck 1959, but without for Whittemore 1997 - alongside toothless B. trifoliolata. The critical role of tooth orientation is not discussed.
Image scans selected from the above treatments with "recurved teeth" leave some doubt as to the adequacy or appropriateness of this broad application of the term.
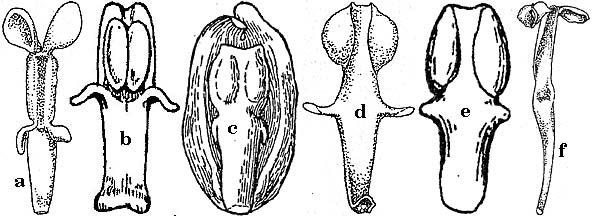
Key: a. B. dictyota (FNA),
b. B. pinnata (Jepson 1925),
c. B. species (Munz & Keck),
d. B. aquifolium (FNA),
e. B. piperiana (Abrams 1944),
f. B. trifoliolata (FNA).
Somewhat less derivative are Hitchcock & Cronquist 1964. Although in their genus description they have "filaments sometimes toothed at the tip" (note that one of their three species is B. nervosa), their specification of B. aquifolium gives:
filaments usually with 2 short, more or less divergent teeth at the tip just below the anthers
The notion of divergence (cf. Gray 1895) would seem to express the symmetrically opposite pairing of the teeth from a common point. The Hitchcock & Cronquist drawing of B. aquifolium might be interpreted as placing this point on the connective, well above the filament.
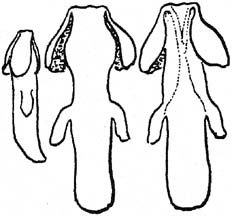
B. aquifolium stamens (adapted from Hitchcock & Cronquist 1964)
Among the West Coast species only B. nervosa is inappendiculate, and it was equally well distinguished by its leaflet venation, lengthy and long persisting bud scales. Distinctive filament structure could be omitted, as was the case in early Jepson 1911 as well as the recent Williams 1993 revision in The Jepson manual : higher plants of California, and numerous other western florae.
In Texas and New Mexico the inappendiculate B. trifoliolata occupies a comparable position, being sufficiently distinguished by its palmately trifoliate leafs. Thus florae of Texas (Correll & Johnston) and New Mexico (Martin & Hutchins 1980), make no mention of filaments.
For Arizona, with one additional trifoliate species, B. harrisoniana, Kearney & Peters 1960 rely on filament structure to distinguish it from B. trifoliolata (p. 320):
filaments conspicuously bidentate, with spurlike teeth